我国的水稻育种经历了矮化育种、优势利用和绿色超级稻培育3次飞跃,其间伴随矮化育种(第一次绿色)、三系杂交稻培育、二系杂交稻培育、亚种间优势利用、理想株型育种和绿色超级稻培育等6个重要历程。育种目标从唯产量是举到高抗、优质和高产并重,育种理念从高产优质逐步提升为“少投入,多产出,保护环境”。水稻功能基因组研究为第二次绿色准备了大量的有重要利用价值的基因,水稻育种正迈向设计育种的新时代。基因组选择技术和转基因技术将为培育“少打农药,少施化肥,节水抗旱,优质高产” 绿色超级稻保驾护航。
本文对我国水稻遗传育种的发展历程进行了概括,指出了各种育种方法和育种技术的优缺点,系统介绍了水稻细胞质雄性不育和光温敏雄性核不育以及籼粳不育的分子机制的研究进展,综述了水稻株型、穗型、粒形和养分高效利用相关的重要功能基因,阐明了产量与开花期联动的关系,凸显了我国水稻基础研究在国际上的重要地位。特别指出,近年来,我国水稻生产方式发生了或正在发生巨大变革,育种理念也要与时俱进。未来,杂交育种技术要与现代育种技术紧密结合,选育水稻品种不仅要满足市场需求,而且更要具备绿色健康的特点,同时还要适应新耕作制度和新耕作方法。
在水稻种植历史中,无论是常规地方品种、现代品种还是杂交组合,产量一直是重要考量目标之一。特别是20世纪中期以来,我国农业生产力低下,为了解决粮食短缺问题,我国育种家唯产量是举,培育出耐大肥大水的品种,依靠大量使用农药化肥,虽然解决了燃眉之急,但也给我国的生态环境造成了破坏,这种生产模式不具有可持续性。21世纪以来,人们生活水平显著提高,我国的水稻生产渐渐地发生了重大变化,育种目标也从单一高产转向优质高抗高产等复合性状目标,绿色农业指日可待。
水稻地方品种基本是高秆类型,耐肥力差,容易倒伏,导致稳产问题。因此,发掘矮秆种质资源,培育矮秆品种变得十分重要。1956年,我国育种家黄耀祥先生以广西农家品种“矮仔占”为材料,选育出“矮仔占4号”,并与高秆品种“广场13”杂交,培育出第一个矮秆籼稻品种“广场矮”。1966年,国际水稻研究所(IRRI)利用我国省地方品种低脚乌尖(Dee-geo-woo-gen)与皮泰(Peta)杂交,育成了半矮化的品种IR8,创造了当时的高产奇迹。我国的矮化育种比国际上整整提前了10年。“广场矮”的培育以及IR8的引进推动了我国水稻育种进入第一次“绿色”时代,水稻亩产从50年代的164千克提高到70年代初的238千克,水稻单产提高了将近45%,实现了水稻产量的第一次飞跃。实际上矮秆品种单株产量略微下降,但是由于其耐密植和抗倒伏,群体产量大幅度提升;同时矮秆品种的耐肥能力强,氮素吸收利用效率低,大量氮肥的施用在提高产量的同时使土壤和环境受到破坏。
优势是指一个物种的不同品种或者物种间的杂交后代的生物量、发育速度和产量的表型值优于两个亲本的现象。我国稻作科学的奠基人丁颖先生曾用人工办法给水稻“去雄”,但实际效果不佳。1964年,湖南安江农校的袁隆平先生开始杂交水稻育种研究,并提出优势利用的设想。1970年,袁隆平先生和他的助手李必湖等人在海南三亚发现了花粉败育的野生稻,花粉败育是由不育细胞质产生(CMS- WA),为杂交水稻雄性不育系的选育打开了突破口。1971年,杂交水稻课题被列为全国协作项目,野败材料被分发到国内水稻科研单位。1973年10月,在苏州召开的水稻科研会议上,袁隆平先生发表《利用“野败”选育“三系”进展》的论文,标志中国籼型杂交水稻三系(不育系、保持系和恢复系)配套成功。同时,江西省萍乡市农业科学研究所的颜龙安先生利用“野败”育成不育系珍汕97A。1981年,福建省三明市农业科学研究所的谢华安先生育成具有抗病性强、配合力高、米质优良的恢复系明恢63。明恢63与珍汕97A配制的强优势组合汕优63产量高,且具有抗病、耐低磷钾、耐高低温、米质较好、适应性广等特点,累计推广6287.7万公顷。三系杂交稻平均产量比一般普通良种增产20%左右,亩产能达到430公斤。
CMS-WA是细胞质基因和核基因互作导致花粉败育类型,属于孢子体雄性不育。可以通过回交的方法保留细胞质基因组而交换核基因组,达到培育不育系的目的。除CMS-WA外,我国水稻育种学家还创制出不同细胞质来源的核质互作雄性不育系。四川农业大学的周开达先生等用西非品种冈比亚卡与朝阳1号、雅安早等杂交和回交,育成冈型不育系冈12朝阳1号A和冈22雅安早A;同时,周开达先生等从Dissi D52/37//矮脚南特群体中选出不育株,育成D型不育系意大利A。意大利A与珍汕97B中变异株杂交,育成D汕A。湖南省农业科学院利用地理远距离品种间杂交组合,育成印尼水田型不育系。冈型、D型和印尼水田型不育特性以及恢、保关系与野败相似。不同育种单位育成了很多孢子体雄性不育类型的不育系,如四川省农业科学院的K型,其胞质不育基因来自于粳稻,恢、保关系与野败相同;安徽省广德县农业科学研究所从江西省引进的矮杆野生稻中发现一株雄性不育株,命名“矮败”,通过核置换育成协青早A,恢、保关系与野败相同,K型和矮败型不育系属于孢子体不育。1972年,武汉大学朱英国先生等以红芒野生稻为母本与莲塘早杂交,选育出红莲型细胞质雄性不育系(HL- CMS),为配子体不育类型。包台型(BT)不育系台中65A引入我国,并经湖南省农业科学院转育成BT型黎明A,与C系统恢复系的配组,使得BT型不育系成为我国粳稻优势利用的主要不育系类型。BT型不育系花粉败育属于配子体不育,不育性没有CMS-WA稳定,高温易自交结实,杂交制种种子不纯。滇型不育系是云南省培育出的适应当地高海拔气候环境的粳稻CMS,属于配子体不育类型。HL-CMS的花粉败育特征为圆败型,而BT- CMS和滇型为染败型。
1973年,湖北沙湖原种场农技员石明松先生发现水稻农垦58的光敏核不育(PGMS)株,并育成了首个光敏核不育系农垦58S。农垦58S在长日高温条件下表现为雄性不育,作为不育系用于杂交水稻制种;在短日低温条件下可育,用于不育系的繁种。PGMS的敏感阶段为幼穗发育时期,主要是从二次枝梗分化到花粉母细胞形成时期,在长日照条件下绒毡层提前降解,缺乏营养供给导致小孢子败育。温度和光照长度在农垦58S中具有补偿效应,高温可以降低临界日照长度,而低温要提高临界日照长度。利用农垦58S光敏不育的特性,突破三系配套恢保关系束缚,开创了“二系”杂交水稻育种的新阶段。另外一类种质例如安农S-1、衡农S-1和5460S,称为水稻温敏核不育(TGMS),温度变化可以导致育性的转换,高温不育,低温可育,而光周期长短对育性转换没有影响。TGMS的诱导阶段在花粉母细胞形成和减数时期,小孢子母细胞不能完成减数,败育的花粉都呈现出皱缩的形态。两系杂交稻由于冲破了恢保关系的束缚,亲本间的遗传差异变大,两系杂交稻平均产量比三系杂交稻具有较大的提高,代表性的品种如两优培九大面积种植能达到亩产630公斤,比三系对照汕优63增产约10%左右。
籼稻和粳稻亚种之间具有更丰富的遗传多样性,杂交组合比籼籼组合具有更强的优势。但是,籼粳杂交种F1不育(或部分可育)限制了籼粳优势的利用。广亲和基因的发现为籼粳亚种间优势利用奠定了理论基础。利用部分粳稻血缘培育的杂交组合例如两优培九、协优9308等表现出很强的优势。直到21世纪以来,利用粳稻不育系与籼粳中间型广亲和恢复系配组配制出籼粳亚种间的杂交品种,如“甬优系列”和“春优系列”组合。这些杂交组合在生产上表现出更强的产量优势。
理想株型是由澳大利亚科学家CM. Donald博士提出,指农作物个体间竞争最小的株型,使每个植株最大限度地获取光照和营养,从而提高群体的收获指数。日本栽培学家松岛省三最早提出高产水稻应该具备“多穗、矮杆、短穗,顶部2、3叶片要短厚直立”的特性。Khush提出少蘖、大穗、茎秆粗壮、株高100~110 cm、叶厚直立、根系发达、晚熟、收获指数高和生产潜力大等特征。我国育种家杨守仁先生等提出高产水稻指标:半矮杆、穗大且直立、分蘖中等,耐肥抗倒、生物量大、谷草比高。袁隆平先生认为超高产杂交水稻在形态上主要特点是上部3片功能叶要长、直、窄、凹、厚,叶面积较大,并且可以两面受光而互不遮蔽,提出库大源足是高产的前提,新株型特征和优势利用相结合是实现超高产水稻育种技术路线年,李家洋先生克隆出理想株型基因IPA1。IPA1植株株型紧凑,茎秆挺直,虽然分蘖能力不强,但成穗率高、穗大、产量潜力大。IPA1的克隆促进了理想株型的育种,培育的理想株型新品种已经表现出巨大的增产潜力。
1999年12月14日,西北农林科技大学的李振声先生、华中农业大学的张启发先生和中国农业科学院作物科学研究所贾继增先生在杭州召开的“农作物资源核心种质构建、重要新基因发掘与有效利用”973项目年会中提出了第2次绿色的10字口号,“少投入,多产出,保护环境”,并提出为绿色准备基因资源。国家973项目“作物高效利用氮磷养分的分子机理”、“主要粮食作物重大病害控制的基础研究”和“害虫爆发成灾的遗传与行为机理”以及农业部948项目“参与全球水稻分子育种计划研究”等,推动了作物营养高效利用和对逆境抗性基因的发掘。2005年和2007年,张启发先生先后两次撰文,提出培育绿色超级稻的构想,主要内容包括“少打农药,少施化肥,节水抗旱,优质高产”,将第二次绿色的基本理念贯穿始终。经过10年的努力,我国选育出一批绿色超级稻品种,全国累计推广约9000万亩。
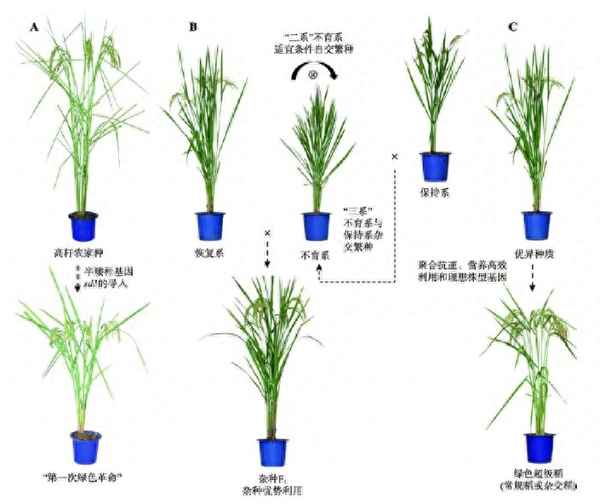
图1 水稻遗传育种经历的3次飞跃。A:半矮杆基因sd1利用以及半矮秆水稻品种培育促成“第一次绿色”;B:不育系和恢复系配制促进优势的利用;C:抗逆、养分高效利用和理想株型等有利基因的发掘促进绿色超级稻的培育。
水稻遗传育种的历程就是一个育种理念变迁的过程。我国水稻遗传育种经历了3次大的飞跃(图1),每次飞跃都离不开重要基因资源的发掘和利用。矮秆基因导致“第一次绿色”,解决了水稻耐肥和抗倒伏的问题;核质互作不育系和光温敏核不育系的培育,促成了优势的利用。第三次飞跃以理想株型塑造为主要技术路线,以绿色超级稻育种为目标,选育高产优质健康新品种/组合,实现第二次绿色。
常规育种包括选择育种、有性杂交育种、物理以及化学诱变育种、离体组织培养育种和细胞杂交育种。常规育种的过程主要是选择合适的亲本,得到分离的群体,利用该育种方法,根据表型从群体后代中选择达到所设定育种目标的个体。这种方法对高产育种效率比较高,但是对稻米品质和非生物逆境的改良效率较低。选择育种是从自然变异中选择优良变异,但是自然变异发生频率低,有价值变异少,育种效率低。诱变育种是通过物理化学处理,增加诱变频率,从大量突变中选择有利突变,由于突变往往是有害的,因此育种效率也低。利用水稻花药培养再生植株,单倍体自然加倍,基因组纯合快,能大量缩短育种历程,但是花药培养严重依赖于基因型,特别是籼稻的花药培养难度较大,因此花药培养育种也受到限制,只有少数单位开展。有性杂交育种是利用不同亲本材料杂交,再通过自交或测交,产生大量的具有丰富表型变异的后代群体,从中选择优良表型的单株。杂交育种充分发挥基因重组的作用,只要亲本间互补性强,杂交育种效率一般比较高,并且很可能育成全新的骨干品种。因此,杂交育种是最主流的水稻育种方法,得到广泛应用。
近30年来水稻功能基因组学的成果为辅助选择育种提供了一系列的功能分子标记。SNP芯片是全基因组选择育种的有效工具,水稻60K SNP芯片的开发和应用为大规模的基因型鉴定提供了便捷的方法。在这些标记辅助选择下,通过回交实现目标性状的定向改良。2017。
jinnian金年会官方网站入口
